
бвШмЩњЬЌЯЕЭГжаЕФжВЮя
ВмНЈЛЊ ЃЌдЌЕРЯШ ЃЌбю Лл ЃЌЛЦ Зв ЃЌжьЭЌБђ ЃЌСКНЈКъ ЃЌжмУЯЯМ ЃЌТолОйЉ ЃЌЮт ЯФ
ЃЈ1. жаЙњЕижЪПЦбЇдКбвШмЕижЪбаОПЫљ/здШЛзЪдДВПЁЂЙуЮїбвШмЖЏСІбЇжиЕуЪЕбщЪв, ЙуЮї Й№Сж 541004ЃЛ2. СЊКЯЙњНЬПЦЮФзщжЏЙњМЪбвШмбаОПжааФ, ЙуЮї Й№Сж 541004ЃЉ
0 в§ бд
бвШмЩњЬЌЯЕЭГЪЧЪмбвШмЛЗОГжЦдМЕФЩњЬЌЯЕЭГЃЌжЇГХЕФбвЪЏЪЧЬМЫсбЮбвЃЌЦфгЩЮоЛњЛЗОГКЭЩњУќЯЕЭГСНИіВПЗжзщГЩЁЃЮоЛњЛЗОГгаСНИіЬиЕуЃКвЛЪЧЬМЫсбЮбвЕФИЛИЦадЃЌГЩЭСЮяжЪЯШЬьВЛзуЃЌЭСШРзЪдДЖЬШБЁЂИЛИЦЦЋМюадЃЛЖўЪЧЕиБэЁЂЕиЯТЫЋВуНсЙЙЃЌЫЎзЪдДвдЕиЯТЫЎзЪдДЮЊжїЁЃЖјЮоЛњЛЗОГЕФСНИіЬиеїгеЗЂЩњУќЯЕЭГЕФСНИіЬиеїЃКвЛЪЧКЕЩњЁЂЪЏЩњКЭЯВИЦЕФбвШмжВБЛЃЛЖўЪЧЮоЙтЁЂГБЪЊЃЌдкЯрЖдКуЮТЬѕМўЯТЕФЕиЯТЩњЮяШК[1-2]ЁЃЬМЫсбЮбвЕФЛЏбЇзщГЩКЭбвШмЕижЪНсЙЙЩюПЬгАЯьбвШмЩњЬЌЯЕЭГЕФЩњВњепЁЂЯћЗбепКЭЗжНтепЃЌгАЯьбјЗжЁЂЫЎЗжЕФЧЈвЦКЭзЊЛЏЃЌОпУїЯдЕФЕижЪгЁМЃ[3-4]ЁЃ
ЩњЬЌЯЕЭГЪЧжИвЛИіЩњЮяШКТфМАЦфжмБпЕижЪЕиРэЛЗОГЯрЛЅзїгУЕФздШЛЯЕЭГЃЌЩњЬЌЯЕЭГгЩЮоЛњЛЗОГЁЂЩњВњепЁЂЯћЗбепКЭЗжНтеп4 ВПЗжзщГЩЃЌЩњЮяШКТфЪЪгІжмБпЛЗОГЖјЩњДцЁЂЗЂг§КЭбнЛЏ[5]ЁЃжВЮяЪЧЩњЬЌЯЕЭГЕФЩњВњепЃЌЖдЮЌГжЩњЬЌЯЕЭГЕФЮяжЪбЛЗЁЂШКТфбнЬцЦ№ЕНжСЙиживЊзїгУЃЌГЄЦкНјЛЏКЭздШЛбЁдёВњЩњЕФбвШмжВБЛЖРОпЬиЩЋ[6]ЁЃБОЮФДгбвШмЩњЬЌЯЕЭГжажВЮяЕФЯВИЦадЁЂЪЏЩњадЁЂКЕЩњадЁЂбјЗжЩњЬЌМЦСПЁЂЖрбљадКЭЬигаадЁЂЩњЬЌЯЕЭГЬМЛуЙІФмЕФаЭЌадЕШЗНУцНјааТлЪіЃЌзюКѓНщЩмСНжжЬигаЕФЬьПгжВЮяШКТфКЭЖДПкжВЮяШКТфЃЌвдЦкЩюШыШЯЪЖбвШмжВБЛЬиЕуЁЂаЮГЩЛњжЦЃЌЖдДрШѕбвШмЩњЬЌЯЕЭГЕФБЃЛЄКЭаоИДЁЂЙІФмЕФЗЂЛгЦ№ЕНДйНјзїгУ[7]ЁЃ
1 бвШмЩњЬЌЯЕЭГжВЮяЕФЯВИЦад
бвШмЩњЬЌЯЕЭГЕФИЛИЦЪєадЃЌЦ№дДгкЬМЫсбЮбвЕФИЛИЦадЃЌЫфШЛВЛЭЌЕижЪЪБДњЕФЬМЫсбЮбвЛЏбЇзщЗжгавЛЖЈВювь(ЦфCaO ЕФКЌСПЮЊ27.3%ЁЋ54.33%ЃЌMgO ЕФКЌСПЮЊ0.49%ЁЋ19.66%ЃЌЫсВЛШмЮяЕФКЌСПЮЊ0.41%ЁЋ10.53%)[8]ЃЌЕЋгыаўЮфбвЁЂЛЈИкбвЯрБШОпгавЛИіЙВЭЌЬиеїЃЌФЧОЭЪЧИЦЁЂУОКЌСПИпЃЌдкЬМЫсбЮбвЩЯдаг§ЕФбвШмЩњЬЌЯЕЭГжаЃЌЮоТлЪЧДѓЦјНЕгъЁЂКгЫЎ-ШЊЫЎ-ЕиЯТЫЎЁЂЭСШРКЭжВЮяЬхжаЃЌЦфИЦКЌСПОљЮЊЙшЫсбЮбвЩЯдаг§ЕФЕиДјадЩњЬЌЯЕЭГРраЭжаЕФМИБЖжСЪЎМИБЖ[9-11]ЁЃ
ИЦдкжВЮяЩњГЄЗЂг§ЕФашЧѓЩЯЮЊДѓСПдЊЫиЃЌЕЋдкЯИАћЫЎЦНЩЯЪєгкЮЂСПдЊЫи[12]ЁЃжВЮяЬхФкИЦЕФКЌСПЮЊ0.1%ЁЋ0.5%ЃЌДѓВПЗжДцдкгкЯИАћБкЩЯЃЌCa2+ЖдЯИАћБкЕФЮШЖЈадЁЂЯИАћЕФЭъећадЃЌвдМАЖдЯИАћЗДгІжаЕФЕїПизїгУЃЌдкжВЮяИіЬхЩњГЄЗЂг§Й§ГЬжаЦ№зХВЛПЩЬцДњЕФзїгУ[13-15]ЁЃ
жВ Юя ЯИ Аћ ЭтЕФCa2+ЪЧmmol·L-1ЫЎ ЦНЃЌЯИ Аћ ФкCa2+дМЪЧμmol·L-1ЫЎЦНЛђИќЕЭЃЌЯИАћФкЕФИЦжївЊЗжВМдквКХнжаЃЌЯИАћжЪжаИЦКЌСПНЯЩйЃЈвдЗРИЦгыСзЫсаЮГЩСзЫсИЦГСЕэЃЉЃЌЦфДЮЪЧЯИАћЦїЃЈЯпСЃЬхЁЂФкжЪЭјКЭвЖТЬЬхЃЉЃЌетаЉИЦЫиДІгкЖЏЬЌЦНКтзДЬЌЃЌвдЕїНкЯИАћЭтЕФРызгЛЗОГгыжЪФЄЕФНсЙЙгыЙІФм[13-17]ЁЃ
ИљОнжВЮяЩњГЄЖдЭСШРИЦжЪЕФвРРЕГЬЖШЃЌПЩЖЈадЕиНЋжВЮяЛЎЗжЮЊ5 РрЃКЂйЪШИЦжВЮяЃЈCalciphileЃЉЃКжЛЗжВМдкИЛКЌЬМЫсИЦЕФЛљжЪЩЯЃЛЂкЯВИЦжВЮяЃЈCalcicoleЃЉЃКдкЖрИЦЭСШРЩЯВХФмЩњГЄе§ГЃЃЌНЯЩйЩњГЄдкЫсадЭСШРЃЛЂлЯгИЦжВЮяЃЈCalcifugeЃЉЃКдкЫсадЭСШРЩњГЄСМКУЃЌЭСжаКЌИЦЩдЖрМДЪмЖОКІзїгУЃЛЂмбЧЯгИЦаджВЮяЃКдкЫсадЭСШРЩЯЩњГЄУЏЪЂЃЌКмЩйМћгкЪЏЛвбвЛђЧПИЦадЭСШРЩЯЃЛЂнжаМфаЭЃЈВЛдёЭСаджВЮяЃЉЃКЖдЭСШРКЌИЦСПЖрЩйУЛгаУїЯдЗДгІ[18-19]ЁЃИљОнЕїВщзЪСЯЕФЯЕЭГећРэЃЌвРОнЁЖжаЙњжВБЛжОЁЗЕФЗжРрддђЃЌдкШКЯЕГпЖШЩЯБржЦжаЙњбвШмжВБЛЗжРрЯЕЭГЃЌПЩЗжЮЊЃК3 ИіжВБЛаЭзщЁЂ13 ИіжВБЛаЭКЭ554 ИіШКЯЕ[7]ЁЃ
ЙѓжнУЏРМздШЛБЃЛЄЧјЪЧбЧШШДјбвШмЩСжБЃЛЄЧјЃЌЪЧбаОПбвШмжВБЛШКТфЬиеїРэЯыЕФвАЭтГЁЫљЃЌЦфЩСжШКТфНсЙЙЮЊГЃТЬТфвЖРЋвЖСжЃЌЦфВЛЭЌгкбЧШШДјГЃТЬРЋвЖСжЃЌвьгкББбЧШШДјКЭбЧШШДјжаГЃТЬТфвЖЛьНЛСж[20]ЃЌВЩМЏбвШмЧјЁЂЗЧбвШмЧјжВЮяЦЗжжИї13 жжЃЌЦфжабвШмЧјгыЗЧбвШмЧјЙВгаЦЗжж7 жжжаМфаЭжВЮяЁЃЭЈЙ§ВтЖЈЦфжВЮявЖЦЌЕФИЦзмСПЁЂВЛЭЌИЦЫиаЮЬЌКЌСПЁЂбЧЯИАћзщЗжИЦКЌСПМАИљМЪЭСЭСШРИЦКЌСПКЭpH ЕШЖрЯюжИБъЃЌНвЪОЯВИЦжВЮяЁЂЯгИЦжВЮяКЭжаМфжВЮяЦЗжжМфЕФЖЈСПВювьадЃЈБэ1ЃЉЁЃ

Бэ1 ЪШИЦжВЮяЁЂЯгИЦжВЮяКЭжаМфжВЮягыИЦЯрЙижИБъЕФЖдБШ[15]Table 1 Comparison of calcium-related indexes among Calciphile, Calcifuge and intermediate plants
ЪШИЦжВЮявЖЦЌИЦжЪКЌСПЦНОљЮЊ1 356.52 mg·kg-1ЃЌБШЯгИЦжВЮяИпГі82.49%ЃЛЪШИЦжВЮявЖЦЌжаИЦвдЙћНКЫсИЦаЮЬЌЮЊжїЃЌЦфКЌСПеМзмИЦСПЕФ27.91%ЁЋ32.82%ЃЌЦфДЮЪЧВнЫсИЦЃЈ23.46%ЁЋ23.96%ЃЉЃЌЯгИЦжВЮявЖЦЌжаИЦжївЊвдВнЫсИЦЮЊжїЃЌЦфКЌСПеМзмИЦСПЕФ33.69%ЁЋ34.34%ЃЌЦфДЮЪЧЙшЫсИЦЃЈ19.35%ЁЋ30.45%ЃЉЃЛЪШИЦжВЮявЖЦЌжаИЦжївЊИГДцдкЯИАћБкЃЌЦфКЌСПеМзмИЦСП59.05%ЁЋ66.54%ЃЌЦфДЮЪЧЯИАћжЪЃЈ23.60%ЁЋ25.76%ЃЉЃЌЯгИЦжВЮявЖЦЌжаИЦжївЊИГДцдкЯИАћжЪжаЃЌЦфКЌСПеМзмИЦСП45.22%ЁЋ46.01%ЃЌЦфДЮЪЧЯИАћБкЃЈ38.07%ЁЋ41.60%ЃЉЁЃ
ЪШИЦжВЮявЖЦЌжаЙћНКЫсИЦЁЂВнЫсИЦЪЧСНДѓжївЊаЮЬЌЃЌЙћНКЫсИЦЪЧЛюадИЦЃЌгаРћгкИЦРызгЕФЧЈвЦЁЂзЊЛЏКЭЮќЪеЃЌдкЬсИпЯИАћБкИеадЃЌЮЌГжЯИАћФЄЕФЮШЖЈадЗНУцОпгаживЊзїгУ[21-22]ЃЌВнЫсИЦЪЧЖшадИЦЃЌПЩдквКАћФкаЮГЩвьОЇЯИАћЃЌвВПЩдкЯИАћМфЯЖаЮГЩеызДЁЂжљзДЕШИїжжаЮзДЕФОЇЬхГСЛ§[23]ЃЌетгыИпИЦбвШмЛЗОГКЭЪШИЦжВЮяЩњРэЬиадЪЧЮЧКЯЕФЃЌгаРћгкжВБЛЮќЪеДѓСПЕФИЦЁЂгаРћгкЕїПиЯИАћФкЭтИЦЫиЕФЦНКтЙиЯЕЁЃЯгИЦаЭжВЮявЖЦЌжажївЊвдВнЫсИЦЁЂЙшЫсИЦЮЊжїЃЌВнЫсИЦЁЂЙшЫсИЦОљЮЊЖшадИЦЃЌФбвдзЊвЦЁЂЮќЪеКЭдйРћгУЃЌетЛђаэОЭЪЧЯгИЦаЭжВЮяЛКНтЙ§СПИЦВњЩњЖОКІЕФЕїНкВпТдЁЃ
ИљОнвбгабаОПНсЙћНвЪОЃЌбвШмжВЮяЖдИпИЦбвШмЛЗОГЕФЪЪгІВпТджївЊАќРЈФЭЪмЃЈШнШЬЃЉЁЂХХГ§КЭзшЖЯЁЃФЭЪмЛњжЦЪЧжВЮяЯИАћМфЯЖДцдкЯрЖдНЯИпЕФгЮРыєШЛљЃЌОпгаНЯИпЕФбєРызгНЛЛЛСПЃЌЪЙЕУжВЮяПЩШнФЩНЯИпКЌСПЕФИЦСП[23]ЃЛХХГ§ЛњжЦгаСНЗНУцЃЌвЛЪЧЬсИпИЦЕїЕААзУИЕФЛюадЃЌШчЯИАћжЪФЄЩЯCa2+-ATPaseЃЈЯймеШ§СзЫсУИЃЉЛюаддіЧПЃЌФмНЋЯИАћжЪжаЕФИЦдЫЫЭГіЯИАћЃЌдкЯИАћБкжаДЂДц[12,24]ЃЌЖўЪЧИЦЭЈЕРЕФЬивьадЃЌаЮГЩУкИЦЯйЬхЁЂЩѕжСЦјПз[25-26]ЃЌНЋЖргрЕФИЦЫиХХГіЯИАћЭт[27-28]ЃЛзшЖЯЛњжЦЪЧжИжВЮяНЋДгЭСШРжаЮќЪеЕФЙ§СПИЦЙЬЖЈдкИљМЪжмЮЇЃЌаЮГЩИЦЛЏИљЃЌЯожЦЦфЯђЕиЩЯВПдЫЪф[20,29]ЁЃ
гЩгкбвШмЩњЬЌЯЕЭГжаЕФИЦПтОоДѓЃЌгАЯьзХбвШмЩњЬЌЯЕЭГЕФЯЕСаЬиеїЁЃЭСШРИЦЖдгаКІжиН№ЪєДцдкозПЙзїгУЃЌЫсГСНЕПЩЛКНтгаКІН№ЪєЕФгАЯьЃЌгаРћгкЮЌГжЩњЬЌЯЕЭГЕФНЁПЕ[30]ЃЛИЦЫиЖдбвШмЩњЬЌЯЕЭГжаЕФбјЗжбЛЗОпгаЧПСвгАЯьЃЌЭСШРИЦЩѕжСБЛШЯЮЊЖдбвШмЩњЬЌЯЕЭГЭЫЛЏЙ§ГЬОпгажИЪОзїгУ[31]ЁЃР§ШчЃЌУРЙњФЯВПбЧЬиРМДѓ(Atlanta)жнPanola ЩНЧјЩСжЩњЬЌЯЕЭГжаЕФИЦУПФъСїЪЇСПЮЊ12.76 kg·ha-1ЃЌвдДЫЫйЖШ80 a КѓЃЌЩСжЩњЬЌЯЕЭГЕФИЦгЊбјНЋЪЇКтЁЃ
2 бвШмЩњЬЌЯЕЭГжажВЮяЕФЪЏЩњад
бвШмЩњЬЌЯЕЭГБЛШЯЮЊЪЧЩњГЄдкЬМЫсбЮбвжЎЩЯЕФЩњЬЌЯЕЭГ[11]ЃЌетвтЮЖзХбвШмжВЮяШКТфЯЕЭГОпгаЗЂДяЁЂЗсИЛЕФЕиЯТЩњЮяСПвджЇГХЕиЩЯЩњЮяЬхЁЃЙѓжнУЏРМздШЛБЃЛЄЧјбвШмжВБЛЕФИљЯЕЩњЮяСПЕїВщНсЙћЯдЪОЃКЕиЩЯЩњЮяСПБШЭЌЮГЖШЕФЗЧбвШмЧјЩСжЕЭЃЌНіЯрЕБгкЮТДјЩСжЧјЃЛЕЋЕиЯТЩњЮяСПдђВЛНіБШЭЌЮГЖШЗЧбвШмЧјЩСжЕФИпЃЌЖјЧвБШЮТДјЁЂШШДјЗЧбвШмЧјЩСжЕФЖМИпЃЌДя57.49ЁЋ58.15 mg·hm-2ЃЌОјЖдСПЪЧЮТДјЁЂШШДјЗЧбвШмЧјЩСжЕФНќ2 БЖ[32]ЁЃ
жВЮяИљЯЕдНЗЂДяЃЌДгЭСШРЁЂбвЪЏЩюВуЮќЪеЕФЫЎЗжКЭбјЗждНЖрЃЌдНФмЪЪгІИЩКЕКЭЦЖёЄЛЗОГ[32-33]ЁЃЕЭЕШжВЮяПЧзДЕивТЪЧГЃМћЕФжГОгдкЬМЫсбЮбвЧГБэВуЕФЯШЗцжВЮяЃЌЫќЪЧдхРрКЭецОњЕФНсКЯЬхЃЌдхРржївЊЩњДцдкзюБэВу1 mmЃЌЭЈЙ§ЙтКЯзїгУЯђЕивТЬсЙЉбјЗжЃЌецОњдђПЩдкЧГБэВу1ЁЋ2 cm ФкЩњДцЃЌМГШЁИќЖрЫЎЗжЃЌЛЅРћЙВЩњЁЃЖјЕивТжГОгПЩЪЙЬМЫсбЮбвЗчЛЏЫйТЪдіЧП24%ЁЋ64%[34]ЃЌЭЌЪБЬсИпбвЪЏЧГБэВуЕФГжЫЎТЪКЭбвЪЏБэУцбјЗжЕФИЛМЏГЬЖШЃЌдкИФдьЮЂЩњОГЕФЭЌЪБЃЌгеЕМИпЕШжВЮяУШЗЂ[34]ЁЃ
ИпЕШжВЮяШКТфОпгаЯрЫЦЕФЙІФмадзДЁЃЭЈЙ§ЖдбЧШШДјЙ№СжбвШмЧј16 ИіЖДбЈЕїВщЃЌПЩЙлВьЕНжВЮяЕФИљЯЕбибвЯЖЛђЪЊШѓЕФбвБкЯТбгДяЪ§ЪЎУзЩюЃЌВЩМЏ81 ИіЖДбЈжВЮяИљЯЕбљБОЃЌОDNA ВтађБэУїИљЯЕбљЦЗЗжЪє38 ИіжВЮяЦЗжжЃЌЦфжага12 жжжВЮяИљЯЕЯТдњЩюЖШДѓгк10 mЁЃЖјЛЦСЌФОЃЈPistacia chinensisBungeЃЉЁЂЮкшъЃЈSapium sebiferum(Linn.) Roxb.ЃЉгЕгазюГЄИљЃЌГЄЖШПЩДя29 m[35-36]ЁЃЩюИљЯЕжВЮяПЩРћгУбвЪЏСбЯЖЫЎЛђЕиЯТКгЫЎЃЌЖјЧГИљЯЕжВЮяжївЊРћгУБэВуДјЫЎКЭЭСШРЫЎЃЛдкЭЌвЛШКТфжажВЮяИљЯЕНсЙЙКЭЗжВМВювьЖдЫЎЗжЮќЪеКЭдйЗжХфОпгаживЊзїгУЃЌДцдкЛЅРћЙВДцЛњжЦ[37]ЃЌДгЖјЮЌГжздШЛЩњЬЌЯЕЭГжаЮяжжЖрбљадКЭШКТфЮШЖЈадЁЃ
бвШмжВЮяЕФЪЏЩњадЛЙБэЯждкбвЪЏРраЭЁЂЙЙдьЬиеїЖджВБЛШКТфНсЙЙзщГЩЕФЧПСвгАЯьЩЯЁЃвђИЦЁЂУОКЌСПЕФВювьадЃЌЬМЫсбЮбвПЩЗжЮЊЪЏЛвбвЁЂАздЦбвСНДѓРрЃЌдкШмНтЗчЛЏЙ§ГЬжаЃЌЬМЫсбЮбв90%вдЩЯЕФЮяжЪЛсЫцЫЎДјзпЃЌВаСєЯТРДаЮГЩЭСШРЕФвЛАудк4%ЁЋ10%ЃЌвђбвЪЏИЦЁЂУОКЭЫсВЛШмЮяКЌСПЕФаЁСПВювьЃЌдкаЮГЩЭСШРЙ§ГЬжаЕУЕНСЫжизщКЭЗХДѓЃЌЖджВЮяШКТфзщГЩВњЩњУїЯдгАЯь[38]ЁЃЙѓжнРѓВЈЯиУЏРМбвШмЩСжЧј4 000 m2ЗЖЮЇФкЙВМЧТМЕНЮЌЙмжВЮя84 ПЦ180 Ъє344 жжЃКДПЛвбвЧјжївЊвдщЪЪїПЦЁЂеСПЦЁЂКњЬвПЦЮЊжїЃЌЪЎИіживЊжЕзюДѓЕФПЦеМзмживЊжЕЕФ60.69%ЃЛДПАздЦбвЧјдђвдщЛПЦЁЂПЧЖЗПЦЁЂщЪЪїПЦКЭЩНмянЧПЦЮЊжїЃЌЪЎИіживЊжЕзюДѓЕФПЦеМзмживЊжЕЕФ66.10%ЃЛВЛДПЬМЫсбЮбвЧјвдЦсЪїПЦЁЂПЧЖЗПЦЁЂеСПЦЮЊжї[39]ЃЈБэ2ЃЉЁЃ
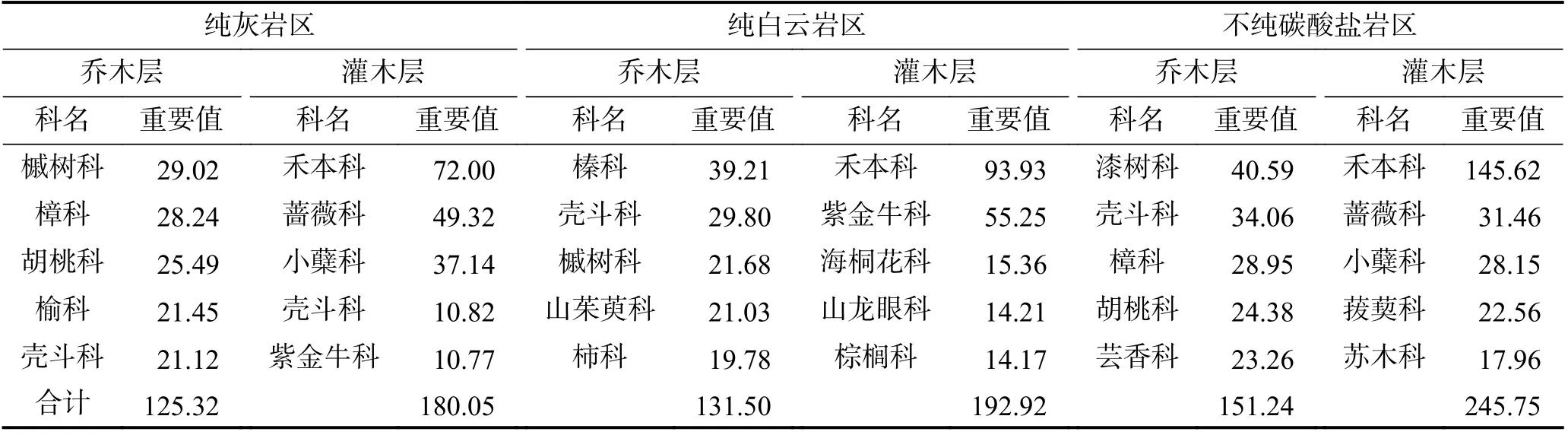
Бэ2 ЙѓжнРѓВЈЯиУЏРМбвШмЩСжЧјШКТфЧЧФОВуКЭЙрФОВужївЊПЦЕФживЊжЕ[39]Table 2 Important values of main families in arbor layer and shrub layer in Maolan karst forest area, Libo county, Guizhou Province
ЪЏЛвбвЕФПѓЮязщЗжЮЊаБСљБпЬхЕФЗННтЪЏЃЌАздЦбвЕФПѓЮязщЗжЪЧе§СљУцЬхЕФАздЦЪЏЃЌЪЏЛвбвИќвзШмНтЗчЛЏЁЂИќвзаЮГЩШмЪДСбЯЖКЭЙмЕРЃЌЧвЯђЕиЯТОпгаИќДѓЕФбгЩьЃЌЧїгкВЛОљдШЗчЛЏЃЌЖјбвЪЏЩюВПЕФШмЪДСбЯЖПЩЮЊФОБОжВЮяИљЯЕЕФЗЂг§ЬсЙЉИќДѓЕФЮяРэПеМфЃЌЛёШЁЩюВуЕиЯТЫЎЃЛАздЦбвПЙШмНтЗчЛЏФмСІНЯЧПЃЌЧїгкОљдШЗчЛЏЃЌЕиБэЕФЭСШРИќЧїОљдШИВИЧЃЌгаРћгкЧГИљЯЕжВЮяЕФЗЂг§ЁЃЙѓжнЪЁЦеЖЈЯиЕїВщЭГМЦНсЙћЯдЪОЃКЪЏЛвбвЮЊжїЕФЕиЧјФОБОжВЮяИВИЧЖШИпЁЂжВБЛЛюЖЏЧП, ЕЋЩњВњСІЫЎЦННЯЕЭЃЛЖјАздЦбвЮЊжїЕФЕиЧјВнБОжВЮяИВИЧЖШИпЁЂжВБЛЛюЖЏШѕЃЌЕЋЩњВњСІЫЎЦННЯИп[40]ЁЃ
3 бвШмЩњЬЌЯЕЭГжажВЮяЕФКЕЩњад
жаЙњФЯЗНбвШмЩњЬЌЯЕЭГЕФИЩКЕЪЧбвШмЫЎЮФЕижЪНсЙЙадИЩКЕЁЃбвШмКЌЫЎНщжЪдкДЙЯђЩЯПЩвРДЮЗЂг§ЮЊЃКбвШмБэВуДјЁЂАќЦјДЙжБЩјСїДјЁЂМОНкВЈЖЏОЖСїДјЁЂБЅЫЎЧБСїДј[41]ЁЃБэВубвШмДјЪЧЕиБэЧПСвбвШмзїгУЙ§ГЬЕФВњЮяЃЌБэВубвШмДјШмЯЖТЪЁЂПзЯЖТЪПЩДя20%ЁЋ60%ЃЌКёЖШЖрдкМИУзжСЪЎМИУз[42-43]ЁЃБэВубвШмДјЕФУмМЏШмЯЖТЪЁЂПзЯЖТЪЃЌМгЩЯЪњОЎЁЂТфЫЎЖДЁЂТЉЖЗЕШЕФЗЂг§ЕМжТНЕгъбИЫйДЙжБШыЩјЁЂЛуШыЕиЯТЃЌКмЩйВњЩњЦТУцОЖСї[44]ЁЃЙ№СжбвШмЧјФмВњЩњЦТУцСїЕФзюЕЭШеНЕгъСПЮЊ64 mmЃЌШЋФъЦТУцОЖСїСПдМеМСїгђФъОЖСїСПЕФ1%ЃЌБэВуДјОЖСїСПдМеМСїгђФъОЖСїСПЕФ8%[45]ЁЃ
ЕиЯТКЌЫЎПеМфвдШмЪДСбЯЖЃЈКЌПзЯЖЃЉЮЊжїЃЌбвШмЖДбЈЛђбвШмЙмЕРжївЊЦ№ДЋЪфзїгУЃЌШчЙѓжнЦеЖЈКѓеЏЕиЯТКгСїгђСљЙШЁЂРЯКкЬЖЕШЫЎЮФеОСїСПЪЕВтзЪСЯНјааЕФЭГВтЗжЮіЯдЪОЃЌСљЙШеОЛуЫЎЗЖЮЇЕФбвШмЙмЕРШнЛ§еМДЂЫЎПеМфЕФ7.47%ЁЋ9.40%ЃЌЖјШмЪДСбЯЖЕФШнЛ§еМ92.53%ЃЛРЯКкЬЖеОЛуЫЎЗЖЮЇЕФбвШмЙмЕРеМДЂЫЎПеМфЕФ7.49%ЁЋ11.60%ЃЌШмЪДСбЯЖЕФШнЛ§еМ88.4%ЁЋ92.5%[46]ЁЃ
ЖјЫЎЪЧЯожЦжВЮяЩњГЄКЭЗжВМЕФзюжївЊвђЫи[31,47]ЁЃЕиЯТЫЎЗсИЛЁЂЕиБэЫЎЖЬШБЃЌГЩЮЊбвШмЧјжВЮядкКЕМОЩњЬЌашЫЎСПВЛзуЕФЕижЪдвђЁЃбвШмЧјЕФжВЮяжївЊЭЈЙ§ЩњЬЌЁЂЩњРэЕФЕїПиРДЪЪгІбвШмИЩКЕЁЃ
жВЮяЭЈЙ§ЖдЩњЬЌадзДЕФЕїПиЃЌвдЬсИпЪЪгІбвШмЧјИЩКЕФмСІЁЃвЛЪЧжВЮяИљЃЈЯЕЃЉЩњЬЌадзДЕФЕїПиЃЌбгГЄИљЃЈЯЕЃЉГЄЖШКЭСПЃЌЛёШЁИќЩюВуЕФЫЎЗж[33]ЃЛЖўЪЧжВЮяФОжЪВПЩњЬЌадзДЕФЕїПиЃЌФОжЪВПжївЊгЩЕМЙмЁЂБЁБкзщжЏКЭЯЫЮЌзщГЩЃЌЕМЙмЪфЫЭЫЎЗжЁЂБЁБкзщжЏДЂДцЫЎЗжЁЂЯЫЮЌЛњаЕжЇГХ[48]ЃЌЫѕаЁЕМЙмжБОЖЁЂЗсИЛБЁБкзщжЏЃЌФмЙЛдкШБЫЎЛЗОГЯТЮЌГжЫЎЗждЫЪфЃЌДгЖјЮЌГжвЖЦЌКЭжІЬѕЕФЩњРэЙІФмзїгУ[49-50]ЃЌЩѕжСгааЉбвШмжВЮяЃЈШчБЁвЖбђЬуМзBauhinia tenuifloraЃЉЕФФОжЪВПЕМЙмОпгаЖРЬиЕФ“ЖўдЊад” ЃЌМДЭЌЪБОпгаДжДѓЕФЕМЙм(ЃО400 μm)КЭжкЖрЯИаЁЕМЙм(ЃМ30 μm) ЃЌвдЪЪгІВЛЭЌЫЎЗжЬѕМўЯТЃЌжВЮяЫЎЗжДЋЪфЁЂРћгУЗНЪН[51]ЃЛШ§ЪЧжВЮявЖЦЌЕФЩњЬЌадзДЕФЕїПиЃЌбвШмжВЮяЕФвЖЦЌЁЂгШЦфЪЧИЩКЕЬѕМўИќЮЊбЯжиЕФЩНЖЅжВЮяЃЌвджВЮявЖЦЌБфаЁЁЂНЧжЪВудіКёЃЈЩѕжСЗЂг§РЏжЪВуЃЉЁЂЦјПзжИЪ§НЕЕЭЃЌЗЂДяЕФБэЦЄУЋЁЂеЄРИзщжЏЕШРДМѕЩйЫЎЗжЩЅЪЇ[52]ЁЃ
ЙуЮїТэЩНХЊРМІЕАБЄЕФЩНЖЅКЭЩНбќЩЯОљЩњГЄЧрИдшнЃЌСНепЕФИпЖШЯрВюНіЮЊМИЪЎУзЃЌЕЋЪЧЦфЗјЩфЁЂЦјЮТЁЂЫЎЗжЛђЪЊЖШЕШЬѕМўНиШЛВЛЭЌЁЃЩНЖЅЬѕМўЖёСгЃЌвђДЫЃЌЩНЖЅжВЮявЖЦЌПэЖШЁЂГЄЖШНіЮЊЩНбќЕФ57.30%ЁЂ54.49%ЃЌвЖЦЌЕФКёЖШЁЂНЧжЪВудіКёдіМг39.70%ЁЂ84.67%ЃЌЦјПзжИЪ§НЕЕЭ22.22%[53]ЃЈБэ3ЃЉЁЃ

Бэ3 ВЛЭЌЕиУВЬѕМўЯТЧрИдшнЕФПЙКЕадЬиеїБШНЯ[53]Table 3 Comparison of drought resistance characteristics of Cyclobalanopsis glauca under different geomorphic conditions
ЙуЮїХЊИкбвШмЗхДдЭнЕиЩСжЧјЃЌФъОљЦјЮТЮЊ22 ЁцЃЌФъНЕЫЎСПЮЊ1 150ЁЋ1 550 mmЃЌбЁдёЭЌвЛжВЮяЦЗжжЦЛЦХ(Sterculia monosperma)ЃЌЖдБШВЛЭЌЩњОГжВЮяШКТфЫЎЗжРћгУаЇТЪЃЌЗжВМгкЭнЕиЕФжВЮяЫЎЗжРћгУаЇТЪЃЈWUEЃЉЮЊ 28.12±16.30 μmol·mol-1ЃЌЩНбќЕФЮЊ44.26±7.04 μmol·mol-1ЁЂЩНЖЅЕФЮЊ46.52±8.60 μmol·mol-1ЃЌДгЭнЕиЕНЩНЖЅЃЌЦЛЦХЫЎЗжРћгУаЇТЪЯджјЬсИпЁЃ4 жжЩНЖЅгХЪЦжВЮяЕФЫЎЗжРћгУаЇТЪБШЭнЕиЕФИп65%[54]ЃЈБэ4ЃЉЁЃ

Бэ4 ЙуЮїХЊИкЗхДдЭнЕиВЛЭЌЩњОГжВЮяШКТфЫЎЗжРћгУаЇТЪ[54]Table 4 Water use efficiency of plant communities in different habitats in Nonggang peak cluster depression, Guangxi
ЭЈЙ§жВЮяЗжУкЩњЮяУИЁЂМЄЫиЕШЃЌЕїПижВЮяЩњРэЙ§ГЬЃЌЬсИпЪЪгІбвШмЧјИЩКЕФмСІЁЃР§ШчЃЌЖдбЧШШДјбвШмЧјГЃМћжВЮяЧрИд(Cyclobalanopsis glaucea)ЁЂдВвЖЮкшъ(Sapium rotundifoliumHemsl. )ЁЂЛ№МЌ(Pyracantha fortuneana(Maxim.) Li)КЭаЁЙћЧОоБ(Rosa cymosa)ЕФбаОПЗЂЯжЃЌЫцзХИЩКЕМгОчЃЌетаЉжВЮяЭЈЙ§Л§РлИЌАБЫсЁЂПЩШмадЬЧЕШгаЛњЩјЭИЕїНкМСЕїПижЪФЄЩјЭИЪЦЃЌЮЌЛЄЯИАћФЄЮШЖЈЃЌМѕЩйЫЎЗжаВЦШЖдЯИАћЕФЩЫКІЃЌЭЌЪБРлЛ§ФЦЁЂМиЕШЮоЛњРызгЕїНквКХнЩјЭИЪЦЃЌЮЌГжЯИАћХђбЙ[55]ЁЃбвШмЧјВЛЭЌбнЬцНзЖЮДњБэаджВЮяђЙФО(Excentrodendron hsienmu(Chun et How) H. T. Chang et R. H.Miau)ЁЂЙЙЪї(Broussonetia papyrifeera)КЭЛ№МЌЕШдкЪЪЖШИЩКЕЬѕМўЯТвРППдіМгЬхФкГЌбѕЛЏЮяЦчЛЏУИЃЈSODЃЉЁЂЙ§бѕЛЏЮяУИ(POD)КЭЙ§бѕЛЏЧтУИ(CAT)ЕФКЌСПЃЌвдЧхГ§ЛюадбѕздгЩЛљНЕЕЭФЄжЌЙ§бѕЛЏГЬЖШЃЌБмУтИЩКЕЖдЦфдьГЩЩЫКІЃЌДяЕНЛКНтжВЮяИЩКЕзДПіЕФФПЕФ[56-57]ЃЌЬсИпбвШмжВЮявЖЦЌЬМЫсєћУИЛюадЃЌНЋЯИАћФкЬМЫсЧтИљРызгзЊЛЏГЩЫЎКЭCO2ЃЌВЙГфИЩКЕв§Ц№ЕФвЖЦЌЫЎЗжВЛзуКЭАћМфCO2МѕЩйЃЌдкЬсИпжВЮяЫЎСІАВШЋЕФЭЌЪБЮЌГжЙтКЯВњЮяЕФКЯГЩЃЌЬсИпбвШмжВЮяЕФИЩКЕЪЪгІад[58]ЁЃ
4 бвШмЩњЬЌЯЕЭГжажВЮяЕФЖрбљад
бвШмЛЗОГЕФЩњОГвьжЪаддкВЛЭЌЕФПеМфГпЖШЩЯОљДцдкЃЌВЛЭЌбвШмОАЙлРраЭЁЂВЛЭЌЕиУВВПЮЛЁЂВЛЭЌЦТЯђЁЂаЁЩњОГЫЎЗжКЭбјЗжЕФВювьадЕШЃЌЩњОГЕФвьжЪадПЩгАЯьжВЮяЖрбљадЕФаЮГЩЃЌЩѕжСГЩЮЊжВЮяЖрбљадЮЌГжЕФжїЕМвђзг[59-60]ЁЃ
бвШмЩњЬЌЯЕЭГжаЕФжВЮяЖрбљадГЃБэЯжЮЊЩйЪєПЦЁЂЙбжжЪєЁЃдЦФЯЮїЫЋАцФЩШШДјбвШмЧјУцЛ§дМЮЊ3 600 km2ЃЌеМЭСЕиУцЛ§ЕФ19%ЃЌКЃАЮЮЊ600ЁЋ1 600 mЃЛбвШмЧјЩСжЕїВщНсЙћЯдЪОЃКгЕгаЮЌЙмЪјжВЮя153 ПЦЁЂ640 ЪєЁЂ1 394 жжЃЌЗжБ№еМЧјЯЕзмПЦЪ§ЕФ77.7%ЁЂЪєЪ§ЕФ56.1%КЭжжЪ§ЕФ37.9%[61]ЁЃ
ЙуЮїХЊИкББШШДјбвШмЩСжЧјУцЛ§100 km2ЕФЩюШыЕїВщНсЙћЯдЪОЃЌЮЌЙмЪјжВБЛга184 ПЦЁЂ810 ЪєЁЂ1 752жж[62-63]ЃЌЗжБ№еМЙуЮїжВЮяЧјЯЕЕФ80.7%ЁЂ72.3%ЁЂ58.7%[6]ЃЛЙуЮїФОТлжабЧШШДјбвШмЩСжЧјжВЮяЧјЯЕЮЌЙмЪјжВБЛга175 ПЦЁЂ528 ЪєЁЂ906 жжЃЌЗжБ№еМЙуЮїжВЮяЧјЯЕЕФ76.8%ЁЂ47.1%ЁЂ30.4%[64]ЃЛЙуЮїбвШмЧјУцЛ§дМ8.2 Эђkm2ЃЌеМЭСЕиУцЛ§ЕФ34.8%ЃЌдчЦкЕїВщНвЪОЙуЮїбвШмжВБЛЕФЧјЯЕгЩ175 ПЦЁЂ662 ЪєЁЂ1 500 жжзщГЩ[65]ЃЌеМ Йу Юї жВ Юя Чј ЯЕЃЈ228 ПЦЁЂ1 120 ЪєЁЂ2 983 жжЃЉЕФ76.8%ЁЂ59.1%ЁЂ50.3%[6]ЁЃ
РѓВЈЯиУЏРМжабЧШШДјбвШмЩСжЧјУцЛ§200 km2ЃЌЮЌЙмЪјжВЮяга154 ПЦЁЂ514 ЪєЁЂ1 203 жж[20],еМЙѓжнЪЁжВЮяЧјЯЕЕФ61.6%ЁЂ33.1%ЁЂ21.1%[66]ЃЛЙѓжнбвШмЧјУцЛ§дМ11.6 Эђkm2ЃЌеМЭСЕиУцЛ§ЕФ61.2%ЃЌвбгаЕїВщНсЙћЯдЪОЙѓжнЙВгаЮЌЙмЪјжВЮя250 ПЦЁЂ1 551 ЪєвдМА5 691 жж[67]ЁЃ
ЙѓжнЁЂЙуЮїЁЂдЦФЯШ§ЪЁЃЈЧјЃЉбвШмЕиЧјЕФЮЌЙмЪјжВЮяЙВга195 ПЦЁЂ1 213 ЪєЁЂ4 287 жжЃЌЦфжаАќРЈаэЖрЬигаЪєКЭЬигажж[61,68-69]ЃЌжВЮяжжЪєБШЮЊ3.8ЃЌЯджјЕЭгкШЋЙњ8.5 ЕФЫЎЦНЃЌЗДгГГібвШмЩњЬЌЯЕЭГжажВЮяжжЪєЕФЖрбљадЁЃ
бвШмЩњЬЌЯЕЭГжажВЮяЖрбљадЕФСэвЛИіживЊЬиеїЪЧЮяжжЕФЬигаадЁЃШчЙћНЋбвШмЧјЩњГЄЗЂг§ЕФжВЮяГЦЮЊбвШмжВЮяЃЌдђбвШмжВЮяЕФзщГЩПЩЗжЮЊШ§жжЃКЬигажжЛђзЈгажжЃЌМДжЛФмдкбвШмЧјЩњДцЃЌдкЗЧбвШмЧјдђЮоЗЈЩњДцЃЛЪЪвЫЛђЪЪгІбвШмЛЗОГЕФЪЪвЫжжЃЌжВЮядкбвШмЧјЩњГЄКмКУЃЌдкЗЧбвШмЧјЩњГЄВЛКУЁЂЕЋФмЩњДцЃЛЫцгіжжЃЌМДжВЮяЦЗжжОпгаКмЧПЕФЪЪгІФмСІЃЌдкбвШмЧјКЭЗЧбвШмЧјОљФмСМКУЩњГЄЁЃ
ИљОнжВЮябЇМвЕФЕїВщЗжРрЃЌЮїЫЋАцФЩбвШмЧјвбгаЕїВщЕФжВБЛЦЗжж1 394 жжЃЌЬигажж141 жжЁЂЪЪвЫжж170 жж[70]ЃЌЗжБ№еМЧјЯЕжВЮяжжЪ§ЕФ10.1%КЭ12.2%ЃЛЙуЮїХЊИкББШШДјбвШмЩСжЧјвбгаЕїВщЕФжВБЛЦЗжж1 752жжЃЌЬигажж278 жжЁЂЪЪвЫжж350 жж[63]ЃЌЗжБ№еМЧјЯЕжВЮяжжЪ§ЕФ15.9%ЁЂ20.0%ЃЛЛЛбджЎЃЌОпгабвШмЬиЩЋЕФжВЮяЦЗжжеМЧјЯЕЦЗжж30%вдЩЯЃЌШчЙћбвШмЩњОГдтгіЦЦЛЕЃЌдђ30%ЕФОпгаЬиЩЋЕФжВЮяЦЗжжЪзЯШЩЅЪЇЃЌЧвЛжИДФбЖШЪЎЗжРЇФбЁЃР§ШчЃЌЙѓжнРѓВЈЯиУЏРМбвШмЩСжБЃЛЄЧјЃЌБЛСаШыIUCN ЮяжжУћТМКЭжаЙњЮяжжКьЩЋУћТМЕФжВЮяжжРрЩцМА27 ПЦЁЂ67 ЪєЁЂ121 жжЃЌдк121 жжжаЃЌБєЮЃжВЮяга83 жжЃЌеМзмЪ§ЕФ68.6%ЃЌАќРЈЙѓжнЩНКЫЬвЃЈCarya KweichowensisЃЉ ЁЂ ЯС вЖ КЌ аІЃЈMichelia angustioblongaЃЉЁЂРѓВЈСіЙћВшЃЈCamellia rubinuricataЃЉЁЂАзЛЈЖЕРМЃЈPaphiopedium emersoniiЃЉЕШ[71]ЁЃ
5 бвШмЩњЬЌЯЕЭГжаСНРрЬигажВЮяШКТф
5.1 бвШмЖДбЈШѕЙтДјжВЮя
бвШмЖДбЈжВЮяЃЌЪЧжИбвШмЖДбЈЩњОГжаздШЛЗжВМЕФжВЮяЁЃИљОнЙтееЧПЖШКЭЭтВПЦјКђгАЯьГЬЖШЃЌЖДбЈПЩБЛЗжЮЊ3 ИіЧјгђЃКЖДПкДјЃЈcave entrance zoneЃЉЁЂШѕЙтДјЃЈcave twilight zoneЃЉКЭКкАЕДјЃЈcave dark zoneЃЉ[72]ЁЃгЩгкКкАЕДјЕФЙтееЧПЖШМИКѕЮЊ0ЃЌНіФмЩњГЄМЋЩйЕФЁЂШБЗІвЖТЬЫиЕФжВЮяКЭвЛаЉецОњЁЂЯИОњЕШЃЌвђДЫбвШмЖДбЈжВЮяДѓЖрЗжВМгкЖДПкДјКЭШѕЙтДјЁЃ
ОнВЛЭъШЋЭГМЦЃЌжаЙњДг2001-2021 ФъМфЖДбЈжВЮяаТжжЗЂБэГЌЙ§СЫ80 жжЃЌЦфжаУшЪізюЖраТЮяжжЕФЧА4ШЫЃЈЕквЛЖЈУћШЫЃЉЗжБ№ЪЧеХРіБјЃЈ23 жжоЇРржВЮяЃЉЁЂЮТЗХЃЈ20 жжжжзгжВЮяЃЉЁЂЭѕЮФВЩЃЈ17 жжжжзгжВЮяЃЉЁЂСѕбнЃЈ11 жжжжзгжВЮяЃЉЁЃ
ЕїВщжаЙњЮїФЯЕиЧјЕФ60 ЖрИібвШмЖДбЈЃЌЗЂЯж83ПЦ186 Ъє418 жжЮЌЙмЪјжВЮяЃЌЦфжа31 жжЃЈ7%ЃЉЮЊЖДбЈЬигажжЁЂ37%ЮЊжаЙњЬигажжЁЃ88%ЕФжжРрЮЊВнБОЁЂ8%ЮЊЙрФОЁЂ4%ЮЊЬйБОЃЌЦфжаЮяжжзюЗсИЛЕФПЦАќРЈЃКнЁТщПЦЃЈ73 жжЃЉЁЂПрмФЬІПЦЃЈ37 жжЃЉЁЂЧяКЃЬФПЦЃЈ22 жжЃЉЁЂЗяЮВоЇПЦЃЈ20 жжЃЉЁЂСлУЋоЇПЦЃЈ20 жжЃЉЃЛзюЗсИЛЕФЪєАќРЈЃКТЅЬнВнЪєЃЈ42 жжЃЉЁЂЧяКЃЬФЪєЃЈ22 жжЃЉЁЂЖњоЇЪєЃЈ19 жжЃЉЁЂБЈДКмФЬІЪєЃЈ19 жжЃЉЁЂРфЫЎЛЈЪєЃЈ13 жжЃЉ[73]ЁЃ
ЙуЮїжВЮябаОПЫљжВЮяЖрбљадбаОПЭХЖгбаОПСЫбвШмЖДбЈДњБэРрШКБЈДКмФЬІЪєжВЮяЖдЕЭЙтЛЗОГЕФЗжзгЪЪгІЛњжЦЃЌЗЂЯжгыЙтКЯзїгУЯрЙиЕФЙтУєЩЋЫиМвзхPHYEЛљвђЕФСНИіЮЛЕуЪмЕНе§ЯђбЁдёЃЌвђЖјЭЦВтИУЙІФмЛљвђдкЦфЪЪгІЖДбЈЛЗОГЕФЙ§ГЬжаПЩФмЦ№ЕНЙиМќзїгУЃЌбвШмЖДбЈЮЊжВЮяЮяжжЕФаЮГЩКЭЪЪгІадНјЛЏЬсЙЉгаРћЬѕМўЕФГЁЫљ[74]ЁЃЖДбЈжВЮявЖЦЌдЊЫиКЌСПЖдЖДбЈЛЗОГЪЧЧїЭЌЪЪгІ[75]ЁЃ
5.2 бвШмЬьПгжВЮяШК
ЬьПгЪЧбвШмЕиЯТКгДѓЬќЖЅАхБРЫњЃЌгЩЬьДАж№НЅбнБфЖјРДЃЌЬьПгЕФЩюЖШЭЈГЃДѓгк100 mЃЌЬьПгЛЗОГЯрЖдЗтБеЃЌЕзВПгыЕиЯТКгЯрСЌЃЌФмИќКУЕиЕїНкЬьПгФкЕФЮТЖШЁЂЪЊЖШЃЌЮЌГжаЁЦјКђЃЌЪЙЦфНЯЩйЪмЭтНчЦјКђБфЛЏИЩШХЃЌЮЊжВЮяЩњДцЁЂЗЂг§ЬсЙЉСМКУЁЂЮШЖЈЕФЛЗОГЃЌФмБЃг§ИќЖрЙ§ШЅвХСєЯТРДЕФжВЮяШКТфГЩЗжЁЃОЕїВщЭГМЦЃЌЙуЮїРжвЕЬьПгжВЮяШКТфжжзгжВЮяга137 ПЦЁЂ445 ЪєЁЂ863 жжЃЌЮТДјПЦКЭШШДјПЦжЎБШЮЊ1ЁУ2.31ЃЌЮТДјЪєКЭШШДјЪєжЎБШЮЊ1ЁУ1.5ЃЛгыПгЭтжВБЛШКТфЃЈ177ПЦ945 Ъє2 406 жжЃЛЮТДјПЦКЭШШДјПЦжЎБШЮЊ1ЁУ2.79ЃЛЮТДјЪєКЭШШДјЪєжЎБШЮЊ1ЁУ2.18ЃЉЯрБШЃЌЖрбљаддкПЦЁЂЪєЁЂжжЫЎЦНЩЯЗжБ№НЕЕЭ22.59%ЁЂ52.91%ЁЂ64.13%ЃЌЕЋПгФкжВЮяШКТфжаЮТДјГЩЗжБШР§ИќИпЃЌАЕЪОзХЫцзХШЋЧђЦјКђБфХЏЃЌЕиБэжВБЛШКТфжаШШДјзщЗжВЛЖЯдіМгЃЌЖјПгФкжВЮяШКТфШШДјзщЗждіМгжЭКѓЁЃетЮЊбаОПЧјгђжВЮяШКТфбнЬцЬсЙЉСЫИќЖрбљБО[76-77]ЁЃ
ЬьПгжВЮяЧјЯЕЕФбнЛЏЯрБШЬьПгЕижЪЗЂг§жЭКѓЃКЬьПгЗЂг§ЧАЦкЃЌЕиЯТДѓЬќПЊЪМБРЫњЃЌПгПкУцЛ§аЁгкПгЕзУцЛ§ЃЌЬьПгаЮзДГЪЕЙжУТЉЖЗзДЃЌЭИШыЕФЙтееЩйЁЂПеЦјГБЪЊЃЌЙтееЧПЖШгЩПгПкЯђПгФкж№НЅЕнМѕЁЃЬьПгФкжВБЛЧјЯЕЦ№дДОРњЬІоКжВЮяНзЖЮЁЂЬйРржВЮяНзЖЮКЭФЭвѕВнБО-ЙрФОНзЖЮЃЛЬьПгЗЂг§жаЦкЃЌЬьПгПкгыЬьПгЕзУцЛ§ЯрНќЃЌДЫЪБЬьПгБЃЮТЁЂБЃЪЊаЇЙћНЯКУЃЌДгЬьПгПкЭИШыЕФЙтеедіЖрЁЂПеЦјГБЪЊЁЂЭСШРМгКёЁЃДЫЪБЬьПгжВЮяЧјЯЕОпгаЧПСвЕФШШДјадЃЌШШДјаджЪЕФЪєЕФБШР§ИпДя70%ЃЌБШЕиБэжВЮяЧјЯЕШШДјЪєЕФБШР§Ип10%ЃЌЯрЗДЃЌЮТДјЪєЕФБШР§БШЬьПгЭтЮТДјЪєЕФБШР§аЁ7%ЃЛЬьПгЗЂг§КѓЦкЃЌЬьПгЭЫЛЏЃЌаќбТВЛЖЯБРТфЃЌПгЕзВЛЖЯБЛЕцЃЌаЮзДГЪТЉЖЗзДЃЌПгПкВПУцЛ§ДѓгкЕзВПУцЛ§ЃЌЬьПгЫљВњЩњЕФЪЊШШаЁЛЗОГЬиеїБфЕУВЛУїЯдЃЌЛЗОГБфЕУКЭЕиБэЧїгквЛжТЁЃДЫЪБЃЌжВБЛШКТфШШДјЪєЕФБШР§ЮЊ65%ЃЌЕЋЪЧШдШЛБШЕиБэжВЮяЧјЯЕШШДјЪєЕФБШР§вЊИп5ИіАйЗжЕуЁЃЖјЮТДјЪєЕФБШР§Щ§ИпЃЌДяЕН25%ЃЌж№НЅНгНќЭтНчжВЮяЧјЯЕжаЮТДјЪєБШР§ЃЌЕЋШдвЊаЁ3 ИіАйЗжЕу[76, 78-79]ЁЃ
6 еЙ Эћ
ЃЈ1ЃЉбЇПЦНЛВцЃЌЭъЩЦбвШмЩњЬЌЯЕЭГРэТл гАЯьЩњЬЌЯЕЭГаЮГЩЁЂбнБфЕФвђЫижаЃЌЦјКђвђЫиЁЂЫЎЮФвђЫиЪЧЫмдьЩњЬЌЯЕЭГГЄЦкЕФЁЂЛКТ§ЕФЧ§ЖЏСІЃЌЖјЕижЪЁЂЕиУВвђЫидђЪЧЩњЬЌЯЕЭГЕУвдДцдкКЭЗЂеЙЕФдиЬхКЭЮяжЪЛљДЁЁЃбвШмЩњЬЌЯЕЭГУїЯдЪмЕижЪЬѕМўЕФжЦдМЃЌжВЮязїЮЊЩњЬЌЯЕЭГЕФГѕМЖЩњВњепЃЌЦфЩњВњСІЙиКѕећИіЩњЬЌЯЕЭГЕФЮяжЪКЭФмСПЙЉИјЁЃжВЮяЩњДцЗЂг§ЕФЧАЬсЬѕМўЪЧОпгазуЙЛЕФЫЎЗжКЭбјЗжЃЌЖјбвШмЧјЪЧШБЭСЩйЫЎЁЂИЛИЦЦЋМюЕФбвШмЛЗОГЃЌЩюШыСЫНтжВЮяЪЪгІбвШмЛЗОГЕФФкдкЛњжЦЃЌДгдЊЫиЩњЮяЕиЧђЛЏбЇбЛЗЕФНЧЖШНвЪОбвЪЏ-ЭСШР-жВЮяМфЫЎЗжЁЂбјЗжЁЂдЊЫиЧЈвЦКЭИЛМЏЙцТЩЃЌВЛНівЊНЋНЙЕуЙизЂбвШмБэВуДјбвЪЏ-ЭСШР-ИљЯЕШкКЯЬхЯЕЖджВЮяЩњВњепЕФжЇГХзїгУЃЌИќвЊМгЩюНвЪОжВЮя-бвЪЏМфЯрЛЅзїгУЫљДјРДЕФЩњРэЁЂЩњЬЌЬиадЃЌетОЭашвЊЩњЬЌЙЄзїепгыЕибЇЙЄзїепУмЧаКЯзїЁЃЖрбЇПЦНЛВцЩјЭИЃЌгШЦфЪЧГфЗжЗЂЛгжаЙњбвШмбаОПЕФЕигђгХЪЦЃЌОЁПьЛёЕУжаЙњВЛЭЌЮГЖШЁЂВЛЭЌКЃАЮжВЮяЖдбвШмЛЗОГЕФЪЪгІЛњжЦКЭЦфЩњРэЩњЬЌЬиадЃЌвдГфЗжРћгУбвШмЩњЬЌЯЕЭГНсЙЙЬиеїЃЌШЯЪЖЭЌвЛЕиЧјжВЮяШКТфбнЬцЙ§ГЬКЭФкдкЙцТЩЃЌгШЦфЪЧЕБЧАШЋЧђЦјКђБфХЏЁЂМЋЖЫЬьЦјЦЕЗЂЕФаЮЪЦЯТЃЌЭъЩЦжаЙњбвШмЩњЬЌЯЕЭГРэТлЁЃ
ЃЈ2ЃЉЭЫЛЏбвШмЩњЬЌЯЕЭГаоИДММЪѕиНашбаЗЂ бвШмЩњЬЌЯЕЭГЪЧДрШѕЕФЩњЬЌЯЕЭГЃЌдкШЫЮЊИЩШХЯТЃЌдк20ЪРМЭФЉжС21 ЪРМЭГѕЃЌжаЙњЮїФЯбвШмЧјГіЯжДѓУцЛ§ЕФЪЏФЎЛЏЯжЯѓЃЌЭСЕиЩњВњСІбЯжиЯТНЕЃЌ“вЛЗНЫЎЭСбјЛюВЛСЫвЛЗНШЫ”ЁЃЮЊЪЕМљ“ТЬЫЎЧрЩНОЭЪЧН№ЩНвјЩН”ЕФЩњЬЌЮФУїРэФюЃЌЭЫЛЏбвШмЩњЬЌЯЕЭГЃЈМЋЖЫБэЯжаЮЪНЪЧжиЖШЪЏФЎЛЏЃЉУцСйСНДѓПЦбЇЮЪЬтЃКвЛЪЧдкЕижЪ-ЦјКђВЛЭЌХфжУЯТЃЌНвЪОЦфжВЮяЩњЬЌаоИДЦПОБММЪѕЕФПЦбЇЛљДЁЃЌвдМАЗТздШЛжВБЛШКТфПьЫйЛжИДШЫЙЄХрг§ММЪѕЕФбаЗЂЁЃЖўЪЧЩИбЁЪЪгІадЁЂЪЪвЫадОМУжВЮяЃЌгШЦфИЛИЦОМУжВЮяЕФХрг§ЃЌЗЂеЙИЛИЦЪГЦЗОМУЃЌНвЪОЦфЖдЕижЪ-ЦјКђВЛЭЌХфжУЯТЕФЩњГЄЁЂЗЂг§КЭбнЬцИќаТЙ§ГЬКЭЛњжЦЃЛбаЗЂЯћГ§еЯАЁЂЬиЩЋЩњЬЌЗЪСЯЁЂбгГЄЦфЩњЬЌОМУ“ЩњУќжмЦк”ЕФЯЕСаММЪѕЃЛбаЗЂ“ЛЅРћЙВгЎ”ЕФЬзжжЁЂМфжжММЪѕЃЌЬНЫїПЩГжајЗЂеЙЭООЖЁЃ
жаЙњЮїФЯбвШмЧјБЃДцгаЙѓжнУЏРМЁЂЙуЮїФОТлКЭХЊИкЁЂдЦФЯЮїЫЋАцФЩЕШдЪМЕФбвШмЩСжБЃЛЄЧјЃЌЦфЮЊЬНЫїДрШѕбвШмЩњЬЌЯЕЭГжаЗЂеЙОпЗТздШЛЬиЩЋЕФЩњЬЌВњвЕЃЌИФЩЦЕБЕиОгУёЩњВњЩњЛюЬѕМўЃЌЪЕМљ“ТЬЫЎЧрЩНОЭЪЧН№ЩНвјЩН”ЕРТЗЃЌЬсЙЉСЫжВЮя“жждДПт”КЭ“ЛљвђПт”ЃЌМјгкбвШмЩњОГЧПСвЕФПеМфвьжЪадЃЌЫЎЭСзЪдДПеМфХфжУЕФВювьадЃЌЛЙгаДѓСПЕФПЦбЇЮЪЬтЁЂММЪѕЮЪЬтиНашНтОіЃЌИќЖрбвШмЧјЩњЬЌОМУаЕїаЭЌЁЂЧвПЩГжајЭООЖиНашЬНЫїЁЃ
жТаЛЃКБОЮФзЋаДЪБМфБШНЯВжДйЃЌзЋаДЙ§ГЬжаЕУЕНЖрЮЛжОЭЌЕРКЯЕФзЈМвЕФжЇГжЃЌгШЦфИааЛЙуЮїжВЮябаОПЫљРюЯШчћЁЂЮЄвуИжЕШРЯЪІЁЂФЯФўЪІЗЖДѓбЇЛЦгёЧхРЯЪІЬсЙЉСЫЯрЙиЕФЫиВФКЭЪ§ОнЁЃ